大家好,又见面了,我是你们的朋友全栈君。如果您正在找激活码,请点击查看最新教程,关注关注公众号 “全栈程序员社区” 获取激活教程,可能之前旧版本教程已经失效.最新Idea2022.1教程亲测有效,一键激活。
Jetbrains全系列IDE使用 1年只要46元 售后保障 童叟无欺
遗传算法原理解析
遗传算法(GA)是一种元启发式自然选择的过程,属于进化算法(EA)大类。遗传算法通常是利用生物启发算子,如变异、交叉和选择来生成高质量的优化和搜索问题的解决方案。
借鉴生物进化理论,遗传算法将问题模拟成一个生物进化过程,通过遗传、交叉、突变、自然选择等操作产生下一代的解,并逐步淘汰适应度函数值低的解,增加适应度函数高的解。这样进化N代后就很有可能会进化出适应度函数值很高的个体。
与遗传算法有关的生物学概念:
(1)染色体(chromosome)
生物是由细胞组成,每一个细胞中都有一套相同的染色体。一条染色体由若干基因(gene) 组成,每个基因控制一种特定的蛋白质,从而决定生物的某种特征。所有染色体合称为基因组(genome)。基因组完全决定了一个生物个体。该个体在微观(基因)层次的表现称为基因型 (genotype),在宏观(特征)层次的表现称为显型 (phenotype)。在简单的遗传算法中,将基因组中的若干条染色体看作一整条染色体。
(2) 个体复制
在复制的过程中,父母的染色体通过交叉(crossover)产生子女的染色体。染色体还可以以一定的小概率变异(mutate)。
遗传算法本质上是一种搜索算法,搜索算法的共同特征为:
- 首先组成一组候选解
- 依据某些适应性条件测算这些候选解的适应度
- 根据适应度保留某些候选解,放弃其他候选解
- 对保留的候选解进行某些操作,生成新的候选解


交叉(Crossover):2条染色体交换部分基因,来构造下一代的2条新的染色体。
交叉前:
00000|011100000000|10000
11100|000001111110|00101
交叉后:
00000|000001111110|10000
11100|011100000000|00101
染色体交叉是以一定的概率发生的,这个概率记为Pc 。
变异(Mutation):在繁殖过程,新产生的染色体中的基因会以一定的概率出错,称为变异。变异发生的概率记为Pm 。
变异前:
000001110000000010000
变异后:
000001110000100010000
适应度函数 ( Fitness Function ):用于评价某个染色体的适应度,用f(x)表示。有时需要区分染色体的适应度函数与问题的目标函数。例如:0-1背包问题的目标函数是所取得物品价值,但将物品价值作为染色体的适应度函数可能并不一定适合。适应度函数与目标函数是正相关的,可对目标函数作一些变形来得到适应度函数。
遗传算法伪代码
基本遗传算法伪代码 /* * Pc:交叉发生的概率 * Pm:变异发生的概率 * M:种群规模 * G:终止进化的代数 * Tf:进化产生的任何一个个体的适应度函数超过Tf,则可以终止进化过程 */ 初始化Pm,Pc,M,G,Tf等参数。随机产生第一代种群Pop do { 计算种群Pop中每一个体的适应度F(i)。 初始化空种群newPop do { 根据适应度以比例选择算法从种群Pop中选出2个个体 if ( random ( 0 , 1 ) < Pc ) { 对2个个体按交叉概率Pc执行交叉操作 } if ( random ( 0 , 1 ) < Pm ) { 对2个个体按变异概率Pm执行变异操作 } 将2个新个体加入种群newPop中 } until ( M个子代被创建 ) 用newPop取代Pop }until ( 任何染色体得分超过Tf, 或繁殖代数超过G )遗传算法优化方法:
(1)精英主义(Elitist Strategy)选择:是基本遗传算法的一种优化。为了防止进化过程中产生的最优解被交叉和变异所破坏,可以将每一代中的最优解原封不动的复制到下一代中。
(2)插入操作:可在3个基本操作的基础上增加一个插入操作。插入操作将染色体中的某个随机的片段移位到另一个随机的位置。
遗传算法实例
计算函数
的最小值,其中每个变量的取值区间都是[-1, +1]。
首先,确定适应度的函数:
function y = my_fitness(population)% population是随机数[0,1]矩阵,下面的操作改变范围为[-1,1]population = 2 * (population - 0.5); y = sum(population.^2, 2); % 行的平方和 遗传算法实现:
function [best_fitness, elite, generation, last_generation] = my_ga( ...
number_of_variables, ... % 求解问题的参数个数
fitness_function, ... % 自定义适应度函数名
population_size, ... % 种群规模(每一代个体数目)
parent_number, ... % 每一代中保持不变的数目(除了变异)
mutation_rate, ... % 变异概率
maximal_generation, ... % 最大演化代数
minimal_cost ... % 最小目标值(函数值越小,则适应度越高)
)
% 累加概率
% 假设 parent_number = 10
% 分子 parent_number:-1:1 用于生成一个数列
% 分母 sum(parent_number:-1:1) 是一个求和结果(一个数)
% 分子 10 9 8 7 6 5 4 3 2 1
% 分母 10+9+...+1=55
% 相除 0.1818 0.1636 0.1455 0.1273 0.1091 0.0909 0.0727 0.0545 0.0364 0.0182
% 累加(cumsum) 0.1818 0.3455 0.4909 0.6182 0.7273 0.8182 0.8909 0.9455 0.9818 1.0000
% 运算结果可以看出: 累加概率函数是一个从0到1增长得越来越慢的函数
cumulative_probabilities = cumsum((parent_number:-1:1) / sum(parent_number:-1:1)); % 1个长度为parent_number的数列
best_fitness = ones(maximal_generation, 1);% 记录每一代最佳适应度,先初始化为1
elite = zeros(maximal_generation, number_of_variables);% 记录每一代的最优解,初始化为0
% 子女数量=种群数量 - 父母数量(父母即每一代中不发生改变的个体)
child_number = population_size - parent_number; % 每一代子女的数目
% population_size 对应矩阵的行,每一行表示1个个体,行数=个体数(种群数量)
% number_of_variables 对应矩阵的列,列数=参数个数(个体特征由这些参数表示)
population = rand(population_size, number_of_variables);% 初始化种群
last_generation = 0; % 记录跳出循环时的代数
for generation = 1 : maximal_generation % 演化循环开始
% feval把数据带入到一个定义好的函数句柄中计算
% 把population矩阵带入fitness_function函数计算
cost = feval(fitness_function, population); % 计算所有个体的适应度(population_size*1的矩阵)
% index记录排序后每个值原来的行数
[cost, index] = sort(cost); % 将适应度函数值从小到大排序
% index(1:parent_number)
% 前parent_number个cost较小的个体在种群population中的行数
% 选出这部分(parent_number个)个体作为父母,其实parent_number对应交叉概率
population = population(index(1:parent_number), :); % 先保留一部分较优的个体
% population矩阵是不断变化的
% cost在经过前面的sort排序后,矩阵已经改变为升序的
% cost(1)即为本代的最佳适应度
best_fitness(generation) = cost(1); % 记录本代的最佳适应度
% population矩阵第一行为本代的最优解(精英)
elite(generation, :) = population(1, :); % 记录本代的最优解(精英)
% 若本代的最优解已足够好,则停止演化
if best_fitness(generation) < minimal_cost;
last_generation = generation;
break;
end
% 交叉变异产生新的种群,染色体交叉开始
for child = 1:2:child_number % 步长为2是因为每一次交叉会产生2个孩子
% cumulative_probabilities 长度为 parent_number
% 从中随机选择2个父母出来 (child+parent_number)
mother = find(cumulative_probabilities > rand, 1); % 选择一个较优秀的母亲
father = find(cumulative_probabilities > rand, 1); % 选择一个较优秀的父亲
% ceil:向上取整
% rand 生成一个随机数
% 即随机选择了一列,这一列的值交换
crossover_point = ceil(rand*number_of_variables); % 随机地确定一个染色体交叉点
% 假如crossover_point=3, number_of_variables=5
% mask1 = 1 1 1 0 0
% mask2 = 0 0 0 1 1
mask1 = [ones(1, crossover_point), zeros(1, number_of_variables - crossover_point)];
mask2 = not(mask1);
% 获取分开的4段染色体
mother_1 = mask1 .* population(mother, :); % 母亲染色体的前部分
mother_2 = mask2 .* population(mother, :); % 母亲染色体的后部分
father_1 = mask1 .* population(father, :); % 父亲染色体的前部分
father_2 = mask2 .* population(father, :); % 父亲染色体的后部分
% 得到下一代
population(parent_number + child, :) = mother_1 + father_2; % 一个孩子
population(parent_number+child+1, :) = mother_2 + father_1; % 另一个孩子
end % 染色体交叉结束
% 染色体变异开始,变异种群
mutation_population = population(2:population_size, :); % 精英不参与变异,从2开始
number_of_elements = (population_size - 1) * number_of_variables; % 全部基因数目
number_of_mutations = ceil(number_of_elements * mutation_rate); % 变异的基因数目(基因总数*变异率)
% rand(1, number_of_mutations) 生成number_of_mutations个随机数(范围0-1)组成的矩阵(1*number_of_mutations)
% 数乘后,矩阵每个元素表示发生改变的基因的位置(元素在矩阵中的一维坐标)
mutation_points = ceil(number_of_elements * rand(1, number_of_mutations)); % 确定要变异的基因
% 被选中的基因都被一个随机数替代,完成变异
mutation_population(mutation_points) = rand(1, number_of_mutations); % 对选中的基因进行变异操作
population(2:population_size, :) = mutation_population; % 发生变异之后的种群
% 染色体变异结束
end % 演化循环结束 函数测试:
clc,clear all,close all;
% 调用 my_ga 进行计算
% 求解问题的参数个数 10
% 自定义适应度函数名 my_fitness
% 种群规模 100
% 每一代中保持不变的数目 50 (即交叉率0.5)
% 变异概率 0.1 (1/10的个体发生变异)
% 最大演化代数 10000 10000代
% 最小目标值 1.0e-6 个体适应度函数值 < 0.000001结束
[best_fitness, elite, generation, last_generation] = my_ga(10, 'my_fitness', 100, 50, 0.1, 10000, 1.0e-6);
% 输出后10行
disp(last_generation);
i_begin = last_generation - 9;
disp(best_fitness(i_begin:last_generation,:));
% disp(best_fitness(9990:10000,:));
% disp(elite(9990:10000,:))
% 不要写成这样,因为GA常常在中间就跳出循环了
% 将elite值转化为问题范围内
my_elite = elite(i_begin:last_generation,:);
my_elite = 2 * (my_elite - 0.5);
disp(my_elite);
% 最佳适应度的演化情况
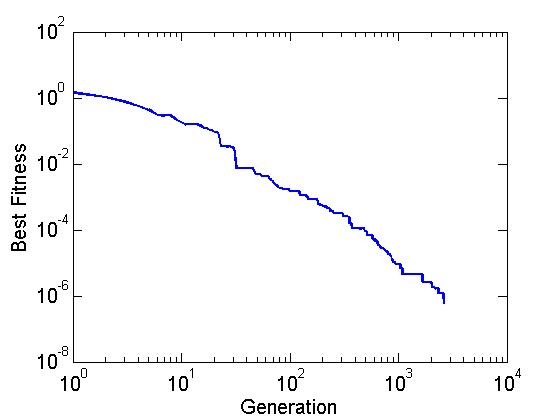
figure
loglog(1:generation, best_fitness(1:generation), 'linewidth',2)
xlabel('Generation','fontsize',15);
ylabel('Best Fitness','fontsize',15);
set(gca,'fontsize',15,'ticklength',get(gca,'ticklength')*2);
% 最优解的演化情况
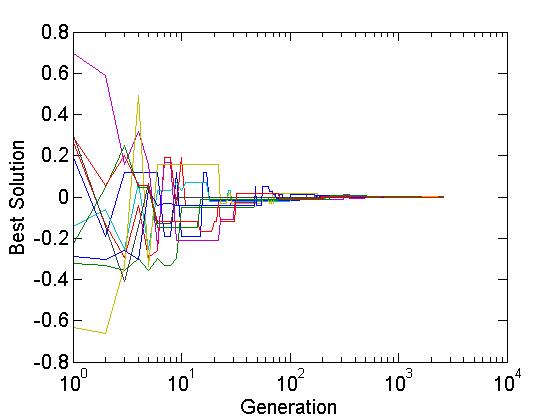
figure
semilogx(1 : generation, 2 * elite(1 : generation, :) - 1)
xlabel('Generation','fontsize',15);
ylabel('Best Solution','fontsize',15);
set(gca,'fontsize',15,'ticklength',get(gca,'ticklength')*2);
参考:
http://blog.jobbole.com/110913/
http://blog.csdn.net/dazhong159/article/details/7908531
http://blog.sciencenet.cn/blog-3102863-1029280.html
https://github.com/Shuai-Xie/genetic-algorithm
https://github.com/yanshengjia/artificial-intelligence/tree/master/genetic-algorithm-for-functional-maximum-problem/src
发布者:全栈程序员-用户IM,转载请注明出处:https://javaforall.cn/194234.html原文链接:https://javaforall.cn
【正版授权,激活自己账号】: Jetbrains全家桶Ide使用,1年售后保障,每天仅需1毛
【官方授权 正版激活】: 官方授权 正版激活 支持Jetbrains家族下所有IDE 使用个人JB账号...
